A few months ago, it was reported that yet another transitional whale fossil was found. Dubbed "Indohyus", it supposedly represented a missing link between land mammals and whales. I was skeptical at first; as I had gotten the impression that the discovers were proposing a brand new theory about the origin of whales and tossing out the old one. After reading an article about the fossil, I realized I had been wrong. The proposal is simply this: The ancestors of whales were small, deer like animals who began as herbivores and later evolved into meat-eaters. Indohyus is NOT thought to be directly related to whales, it is only a 'cousin' which did not change as much as Pakicetus. Here is an easy way to see it:
Pakicetus (Evolves a lot)
\
Common Ancestor
/
Indohyus (Evolves a little)
Click Here for the Scientific American Article.
Thursday, March 27, 2008
Wednesday, March 26, 2008
Fastest Evolving Creature is 'Living Dinosaur'
Livescience reports a very cool finding:
Scientists have pinned down the fastest-known evolving animal — a "living dinosaur" called a tuatara.
The tuatara, Sphendon punctatus, resembles a lizard and is found only in New Zealand. It is the only surviving member of a reptilian order Sphehodontia that lived alongside early dinosaurs and separated from other reptiles 200 million years ago in the Upper Triassic period.
To make the estimate of evolutionary speed, researchers recovered DNA sequences from the bones of ancient tuatara. The team found that although tuatara have remained largely unchanged physically over very long periods of evolution, they are evolving — at a DNA level — faster than any other animal yet examined.The results will be detailed in the March issue of the journal Trends in Genetics.
"What we found is that the tuatara has the highest molecular evolutionary rate that anyone has measured," said researcher David Lambert from the Allan Wilson Centre for Molecular Ecology and Evolution in New Zealand.
Many scientists have thought that molecular evolution would be fastest in animals whose physical form, or morphology, also evolved swiftly. The tuatara finding suggests otherwise, that there is no relationship between the two rates.
"Of course we would have expected that the tuatara, which does everything slowly — they grow slowly, reproduce slowly and have a very slow metabolism — would have evolved slowly," Lambert said. "In fact, at the DNA level, they evolve extremely quickly."
The rate of evolution for Adélie penguins, which Lambert and his team have studied in the Antarctic for many years, is slightly slower than that of the tuatara. The tuatara rate is significantly faster than for other animals that have been studied, including the lion, ox, horse and the now-extinct cave bear.
Lambert says the finding will be helpful in terms of future study and conservation of the tuatara, and the team now hopes to extend the work to look at the evolution of other animal species.
“We want to go on and measure the rate of molecular evolution for humans, as well as doing more work with moa and Antarctic fish to see if rates of DNA change are uncoupled in these species," Lambert said. "There are human mummies in the Andes and some very good samples in Siberia where we have some collaborators, so we are hopeful we will be able to measure the rate of human evolution in these animals too.”
Click here to go to the article.
Scientists have pinned down the fastest-known evolving animal — a "living dinosaur" called a tuatara.
The tuatara, Sphendon punctatus, resembles a lizard and is found only in New Zealand. It is the only surviving member of a reptilian order Sphehodontia that lived alongside early dinosaurs and separated from other reptiles 200 million years ago in the Upper Triassic period.
To make the estimate of evolutionary speed, researchers recovered DNA sequences from the bones of ancient tuatara. The team found that although tuatara have remained largely unchanged physically over very long periods of evolution, they are evolving — at a DNA level — faster than any other animal yet examined.The results will be detailed in the March issue of the journal Trends in Genetics.
{kind=link}
"What we found is that the tuatara has the highest molecular evolutionary rate that anyone has measured," said researcher David Lambert from the Allan Wilson Centre for Molecular Ecology and Evolution in New Zealand.
Many scientists have thought that molecular evolution would be fastest in animals whose physical form, or morphology, also evolved swiftly. The tuatara finding suggests otherwise, that there is no relationship between the two rates.
"Of course we would have expected that the tuatara, which does everything slowly — they grow slowly, reproduce slowly and have a very slow metabolism — would have evolved slowly," Lambert said. "In fact, at the DNA level, they evolve extremely quickly."
The rate of evolution for Adélie penguins, which Lambert and his team have studied in the Antarctic for many years, is slightly slower than that of the tuatara. The tuatara rate is significantly faster than for other animals that have been studied, including the lion, ox, horse and the now-extinct cave bear.
Lambert says the finding will be helpful in terms of future study and conservation of the tuatara, and the team now hopes to extend the work to look at the evolution of other animal species.
“We want to go on and measure the rate of molecular evolution for humans, as well as doing more work with moa and Antarctic fish to see if rates of DNA change are uncoupled in these species," Lambert said. "There are human mummies in the Andes and some very good samples in Siberia where we have some collaborators, so we are hopeful we will be able to measure the rate of human evolution in these animals too.”
Click here to go to the article.
Sunday, March 23, 2008
Expelled
Richard Dawkins has written an excellent article about the new movie 'Expelled' (which he actually went and saw):
Lying for Jesus
Lying for Jesus
Back from Break
Hi Everyone! I haven't made any posts here lately, mainly because I was on Spring Break (I think everyone else was as well). Don't worry, I have more coming: I am planning to do an article about Darwin and Hitler, as well as a little something on the origin of life (again) among other things. Anyway, I made two posts here lately that I don't think a lot of people saw, but I felt like it is among some of my best work, so check it out:
Did Dinosaurs Evolve Into Birds? - Not all scientists think so, and NOT just mean creationists. There is a small group of scientists who believe that the bird lineage split off from reptiles much earlier. I present my opinion on the subject, along with a lot of helpful information.
The Molecular Clock - What it is, and a simple explanation of how it works.
Did Dinosaurs Evolve Into Birds? - Not all scientists think so, and NOT just mean creationists. There is a small group of scientists who believe that the bird lineage split off from reptiles much earlier. I present my opinion on the subject, along with a lot of helpful information.
The Molecular Clock - What it is, and a simple explanation of how it works.
Monday, March 17, 2008
Fighting the Creationists on Youtube
Lately my youtube channel has been subject to two idiotic creationist responses, one of them about abiogenesis and another about my "Proof of Evolution" video.
The guy responding to my abiogenesis video calls himself "Young Apologetic" and is clearly following in the footsteps of other apologists by misquoting prominent scientists and engaging in special pleading. He quotes Francis Crick out of context, and also criticizes Crick's panspermia hypothesis. I, by the way, agree with him that the panspermia hypothesis explains nothing, and that there is no evidence for it. The same applies to intelligent design, and I do not think that an eternal god is a better explanation for the origin of life than a never ending chain of aliens (Panspermia proposes that either the alien's biochemistry was far simpler than ours, or that they were created by another alien civilization which was created by another alien civilization, ad infinitum). Moving on, I think that Wikipedia's entry on Crick summed it up the best:
The conservative political analyst Mark Steyn published an obituary of Crick and attempted a deconstruction of Crick's scientific motivations.[57] Steyn characterized Crick as a militant atheist and asserted that it was his atheism that "drove" Crick to move beyond conventional molecular biology towards speculative topics such as panspermia. Steyn described the theory of directed panspermia as amounting to, "gods in the skies who fertilize the earth and then retreat to the heavens beyond our reach." Steyn categorized Crick’s ideas on directed panspermia as a result of "hyper-rationalism" that, "lead him round to embracing a belief in a celestial creator of human life, indeed a deus ex machina."
To discuss the second video: A girl who appears about ten years old responded to my "Proof of Evolution" video. She may be cute, but the indoctrination and poor reasoning that she demonstrates is anything but. I shouldn't be hard on her, she's young, and as she gets older she will question what she was taught more. Here is how I responded to her:
Hi, I saw you made a video response to "Proof of Evolution". You raise a few points that I think are worth addressing: Where does everything ultimately come from? (you used the example of dust, if that's where everything came from, where did it come from?) Evolution presumes the existence of the universe and life. It simply does not address the questions of life's or the universe's origin. As evolutionist I cannot answer those questions. However, as an atheist and as a person who has just as much wonder about the universe as you do, let me offer you the alternative: http://www.infidels.org/library/modern/mark_vuletic/vacuum.html You also ask what the point is in believing in evolution, or disbelieving in god, since you have nothing to lose by believing and everything to lose by disbelief. Aren't you assuming that god favors blind belief over a rational, well thought out conclusion? This argument is known as Pascal's wager and plenty of philosophers have stepped up to show what a bad argument it is:
http://en.wikipedia.org/wiki/Pascal's_Wager#Atheist.27s_Wager And about the poop, you mentioned it but I don't remember what you asked about it. It is buried in a rock layer that most creationists think was deposited by the flood. Why didn't it dissolve as the rushing waters were burying it? I suppose you could argue that it was already buried, or fossilized, but if it was then it should be underneath the supposed "flood" rock layers, not in it. How would it dissolve the dirt and sediment that the fossil was buried in without dissolving it? Thanks, Ryan
The guy responding to my abiogenesis video calls himself "Young Apologetic" and is clearly following in the footsteps of other apologists by misquoting prominent scientists and engaging in special pleading. He quotes Francis Crick out of context, and also criticizes Crick's panspermia hypothesis. I, by the way, agree with him that the panspermia hypothesis explains nothing, and that there is no evidence for it. The same applies to intelligent design, and I do not think that an eternal god is a better explanation for the origin of life than a never ending chain of aliens (Panspermia proposes that either the alien's biochemistry was far simpler than ours, or that they were created by another alien civilization which was created by another alien civilization, ad infinitum). Moving on, I think that Wikipedia's entry on Crick summed it up the best:
The conservative political analyst Mark Steyn published an obituary of Crick and attempted a deconstruction of Crick's scientific motivations.[57] Steyn characterized Crick as a militant atheist and asserted that it was his atheism that "drove" Crick to move beyond conventional molecular biology towards speculative topics such as panspermia. Steyn described the theory of directed panspermia as amounting to, "gods in the skies who fertilize the earth and then retreat to the heavens beyond our reach." Steyn categorized Crick’s ideas on directed panspermia as a result of "hyper-rationalism" that, "lead him round to embracing a belief in a celestial creator of human life, indeed a deus ex machina."
Steyn's critique of Crick ignored the fact that Crick never held a belief in panspermia. Crick explored the hypothesis that it might be possible for life forms to be moved from one planet to another. What "drove" Crick towards speculation about directed panspermia was the difficulty of imagining how a complex system like a cell could arise under pre-biotic conditions from non-living chemical components. After ribozymes were discovered, Crick became much less interested in panspermia because it was then much easier to imagine the pre-biotic origins of life as being made possible by some set of simple self-replicating polymers.[50]
To discuss the second video: A girl who appears about ten years old responded to my "Proof of Evolution" video. She may be cute, but the indoctrination and poor reasoning that she demonstrates is anything but. I shouldn't be hard on her, she's young, and as she gets older she will question what she was taught more. Here is how I responded to her:
Hi, I saw you made a video response to "Proof of Evolution". You raise a few points that I think are worth addressing: Where does everything ultimately come from? (you used the example of dust, if that's where everything came from, where did it come from?) Evolution presumes the existence of the universe and life. It simply does not address the questions of life's or the universe's origin. As evolutionist I cannot answer those questions. However, as an atheist and as a person who has just as much wonder about the universe as you do, let me offer you the alternative: http://www.infidels.org/library/modern/mark_vuletic/vacuum.html You also ask what the point is in believing in evolution, or disbelieving in god, since you have nothing to lose by believing and everything to lose by disbelief. Aren't you assuming that god favors blind belief over a rational, well thought out conclusion? This argument is known as Pascal's wager and plenty of philosophers have stepped up to show what a bad argument it is:
http://en.wikipedia.org/wiki/Pascal's_Wager#Atheist.27s_Wager And about the poop, you mentioned it but I don't remember what you asked about it. It is buried in a rock layer that most creationists think was deposited by the flood. Why didn't it dissolve as the rushing waters were burying it? I suppose you could argue that it was already buried, or fossilized, but if it was then it should be underneath the supposed "flood" rock layers, not in it. How would it dissolve the dirt and sediment that the fossil was buried in without dissolving it? Thanks, Ryan
Friday, March 14, 2008
Did Dinosaurs Evolve into Birds?
Some of you may have heard of a controversy among scientists: Did birds evolve from birds, or did they evolve from an older reptilian ancestor?
Creationists, of course, have seized this argument and tried to somehow contort disagreement in a single area into a way to show that evolution as a whole is unstable (keep in mind that birds don't fossilize well, so this leaves scientists a lot of room for imagination and speculation. Since different folks have different speculations, a couple of different ideas have been left to duke it out).
I want to state in advance that I am not a scientist, and there may very well be more to this than I know. Most scientists support the view that I will present, but very smart (and very educated) people disagree with the most prevalent view of bird ancestry. There are many arguments launched toward the dinobird theory: A few scientists have claimed that the 'downy feathers' belonging to the dinosaur Sinosauropteryx were really just collagen fibers (these resemble hair and have been known to form on fossils of sharks); there are lots of scientists who think that flight could evolve only by graceful glides from the trees, not hopping up and down on the ground the way theropod dinosaurs must have done.
The first claim that is usually presented against dino bird ancestry is this:
"Most scientists believe that birds descended from theropod dinosaurs. But theropod dinosaurs have lost digits IV and V from a primitively five-fingered hand, leaving them with digits I, II, and III. Birds, on the other hand, have digits II, III, and IV, having lost digits I and V. It is almost impossible for the two groups to be closely related with such a significant anatomical difference."
PZ Myers wrote a wonderful article that explains why this argument is wrong:
Anatomists initially assigned digit numbers I, II, and III to bird limbs on the basis of their form, but later had to revise that to II, III, and IV on the basis of embryology. Dinosaur digits are assigned numbers I, II, and III on the basis of their adult form (which is admittedly much less ambiguous than adult bird digits!)…but what about their embryology? If we had access to information about expression of molecular markers and early condensations in the dinosaur limb, would we have to revise their digit numbers?
Another argument against birds evolving from dinos is that it requires a ground up origin of flight. This is viewed as being less plausible because in order to fly the dinosaurs would have had to work against the force of gravity (The alternative is that the ancestors of birds glided down from trees). This argument is easily answered by simply viewing the latest theories on the evolution of flight (See chapter 3 of "The Four Winged Dinosaur").
In order to find out more about the subject, I emailed Paleontologist Greg Paul:
Hi Greg, I have two questions about bird evolution:
1) What about Dr. Alan Feduccia's claim that bird teeth are not homologous to theropod teeth? (Indicating they are unrelated)
2) Does Sinosauropteryx really have "protofeathers" or is this simply collagen fiber?
Thanks and I hope to hear your response soon,
Ryan
He responded:
Hi Ryan!
Feduccia & company used to argue that dromaeosaurs could not be bird relatives because the formers teeth were too different from those of toothed birds. Because dromaeosaurs are now known to have well developed feathers, Feduccia etc. now argue that dromaeosaurs are birds, not dinosaurs.
New small, short armed theropods from the Yixian show that the feathers formed a soft tuft at the tip of the tail, and came off the upper foot. It is not possible for those to be collagen fibers.
(By the way, velociraptors are dromaeosaurs and they had feathers, as well as others)
One final argument that is often brought up is that it would be impossible for the avian lung system to evolve. To counter that one, simply take a look at how similar the avian and dinosaurian air systems are.
Creationists, of course, have seized this argument and tried to somehow contort disagreement in a single area into a way to show that evolution as a whole is unstable (keep in mind that birds don't fossilize well, so this leaves scientists a lot of room for imagination and speculation. Since different folks have different speculations, a couple of different ideas have been left to duke it out).
I want to state in advance that I am not a scientist, and there may very well be more to this than I know. Most scientists support the view that I will present, but very smart (and very educated) people disagree with the most prevalent view of bird ancestry. There are many arguments launched toward the dinobird theory: A few scientists have claimed that the 'downy feathers' belonging to the dinosaur Sinosauropteryx were really just collagen fibers (these resemble hair and have been known to form on fossils of sharks); there are lots of scientists who think that flight could evolve only by graceful glides from the trees, not hopping up and down on the ground the way theropod dinosaurs must have done.
The first claim that is usually presented against dino bird ancestry is this:
"Most scientists believe that birds descended from theropod dinosaurs. But theropod dinosaurs have lost digits IV and V from a primitively five-fingered hand, leaving them with digits I, II, and III. Birds, on the other hand, have digits II, III, and IV, having lost digits I and V. It is almost impossible for the two groups to be closely related with such a significant anatomical difference."
PZ Myers wrote a wonderful article that explains why this argument is wrong:
Anatomists initially assigned digit numbers I, II, and III to bird limbs on the basis of their form, but later had to revise that to II, III, and IV on the basis of embryology. Dinosaur digits are assigned numbers I, II, and III on the basis of their adult form (which is admittedly much less ambiguous than adult bird digits!)…but what about their embryology? If we had access to information about expression of molecular markers and early condensations in the dinosaur limb, would we have to revise their digit numbers?
Another argument against birds evolving from dinos is that it requires a ground up origin of flight. This is viewed as being less plausible because in order to fly the dinosaurs would have had to work against the force of gravity (The alternative is that the ancestors of birds glided down from trees). This argument is easily answered by simply viewing the latest theories on the evolution of flight (See chapter 3 of "The Four Winged Dinosaur").
In order to find out more about the subject, I emailed Paleontologist Greg Paul:
Hi Greg, I have two questions about bird evolution:
1) What about Dr. Alan Feduccia's claim that bird teeth are not homologous to theropod teeth? (Indicating they are unrelated)
2) Does Sinosauropteryx really have "protofeathers" or is this simply collagen fiber?
Thanks and I hope to hear your response soon,
Ryan
He responded:
Hi Ryan!
Feduccia & company used to argue that dromaeosaurs could not be bird relatives because the formers teeth were too different from those of toothed birds. Because dromaeosaurs are now known to have well developed feathers, Feduccia etc. now argue that dromaeosaurs are birds, not dinosaurs.
New small, short armed theropods from the Yixian show that the feathers formed a soft tuft at the tip of the tail, and came off the upper foot. It is not possible for those to be collagen fibers.
(By the way, velociraptors are dromaeosaurs and they had feathers, as well as others)
One final argument that is often brought up is that it would be impossible for the avian lung system to evolve. To counter that one, simply take a look at how similar the avian and dinosaurian air systems are.
Thursday, March 13, 2008
Dinosaurs with Feathers
You may remember the transitional feather fossils that I posted on a while back. Well, NatGeo did an excellent and informative write up on the find. View it here.
Here are some other dino-bird finds:
T. Rex Cousin Had Feathers
A Giant Turkey-Like Dinosaur
The '7 Foot Turkey'
Velociraptor Had Feathers
Microraptor: The Four Winged Dinosaur
NOVA: The Four Winged Dinosaur
Bird Fossil Transitions
Here are some other dino-bird finds:
T. Rex Cousin Had Feathers
A Giant Turkey-Like Dinosaur
The '7 Foot Turkey'
Velociraptor Had Feathers
Microraptor: The Four Winged Dinosaur
NOVA: The Four Winged Dinosaur
Bird Fossil Transitions
Tuesday, March 11, 2008
The Molecular Clock
Yesterday's post discussed the method of dating by genetics. I think it is important to note that the 'molecular clock' is not foolproof, and is based on assumptions. The molecular clock was derived by estimating when two lineages split in the fossil record (via radiometric dating), and comparing the differences in genetics to come up with a rate of genetic change. Click here for Berkeley University's explanation. But this should not be looked at as precise, because it is based on the following assumptions:
Changing generation times (A mutation generally becomes fixed only from one generation to another. The shorter this timespan is, the more mutations can become fixed)
Population size (Apart from effects of small population size, genetic diversity will "bottom out" as populations become larger as the fitness advantage of any one mutation becomes smaller)
Species-specific differences (due to differing metabolism, ecology, evolutionary history,...)
Evolving functions of the encoded protein (can be ameliorated by utilizing non-coding DNA sequences or emphasizing silent mutations)
Changes in the intensity of natural selection
I thought this was worth mentioning.
Changing generation times (A mutation generally becomes fixed only from one generation to another. The shorter this timespan is, the more mutations can become fixed)
Population size (Apart from effects of small population size, genetic diversity will "bottom out" as populations become larger as the fitness advantage of any one mutation becomes smaller)
Species-specific differences (due to differing metabolism, ecology, evolutionary history,...)
Evolving functions of the encoded protein (can be ameliorated by utilizing non-coding DNA sequences or emphasizing silent mutations)
Changes in the intensity of natural selection
I thought this was worth mentioning.
Sunday, March 9, 2008
The Genome: An Outsider's View
Carl Zimmer wrote an excellent article, and I thought I'd repost it:
The Buddha once told a story about a king who ordered a group of blind men to be presented with an elephant. Each man touched a different part of the animal. The king then asked them what an elephant is like.
The blind men who touched the elephant's head replied, “An elephant, your majesty, is just like a water jar.” The blind men who touched its ear said, “An elephant, your majesty, is just like a winnowing basket.” The blind men who touched its tusk declared, “An elephant, your majesty, is just like a plowshare.” The ones who touched the trunk replied, “An elephant, your majesty, is just like a plow pole.” The blind men who touched the body replied, “An elephant, your majesty, is just like a storeroom.” The blind men who touched the foot replied, “An elephant, your majesty, is just like a post.” The blind men who touched the hindquarters replied, “An elephant, your majesty, is just like a mortar.” The blind men who touched the tail replied, “An elephant, your majesty, is just like a pestle.” And the blind men who touched the tuft at the end of the tail replied, “An elephant, your majesty, is just like a broom.”
The blind men fell into a fistfight, shouting, “An elephant is like this, an elephant is not like that! An elephant is not like this, an elephant is like that!” [1]
I am a science writer, and my chief passion is biology. I spend time with biologists of all stripes—computational biologists, paleontologists, biochemists, ecologists, and all the rest. It is a marvelous privilege. But there are times, I must confess, when I feel like I am watching a blind fistfight.
One of the first bouts I witnessed took place in the late 1990s, when I was researching the origin of whales. Whales descend from terrestrial mammals, and made the transition to water between about 50 million and 40 million years ago. In the 1990s paleontologists began unearthing a series of spectacular fossils documenting that transition, including whales with full-blown legs. Functional morphologists joined forces to investigate the transition, studying swimming otters to understand how proto-whales might have moved through the water. I spent a lot of time with scientists such as these. They are naturalists. They have to know a lot of natural history. They have to memorize the details of many species, to understand how the physiology, behavior, morphology, and ecology of each animal add up into an integrated whole. Yet these naturalists also know that they only have a slippery grasp on all of that embodied complexity.
I put what I learned from those naturalists into my first book, At the Water's Edge [2]. As I was finishing up my manuscript, I began coming across papers in which scientists were taking a radically different approach to the question of whale origins: they were comparing the DNA of whales to that of other mammals.
At the time, molecular phylogenies were still a novelty. The computational methods for calculating them were relatively new, and scientists could only use them to compare a few gene sequences of whales and other mammals. But the conclusions from these few studies were the same: the closest living cousins of whales are hippos.
This may not seem like a big deal. It certainly didn't seem to bother the scientists who carried out the studies. They were just analyzing digital code, abstracted from the animals that carried it. The results spoke for themselves. Nevertheless, they gave paleontologists and mammalian systematists conniptions.
Here's why. Hippos are artiodactyls (also known as even-toed ungulates). Other artiodactyls include cows, camels, and pigs, and goats. Zoologists have long recognized a number of anatomical features that unite artiodactyls in their own group, distinct from other hoofed mammals such as horses and rhinos. One of the most obvious hallmarks was a bone in the ankle, the astragalus. The artiodactyl astragalus has a unique double-pulley shape that allows the artiodactyl hoof to swing back and forth in a distinctive way.
In the 1990s paleontologists found a number of spectacular early whale fossils, but they had yet to find bones from the whale ankle. The bones and teeth they did find suggested that the closest relatives of whales were an extinct group of mammals called mesonychids. Mesonychids were hoofed mammals, but they did not have a double-pulley astragalus. Therefore, paleontologists concluded, they were probably not artiodactyls. And if mesonychids were not artiodactyls, then whales could not be either. And that meant that the whale–hippo link had to be wrong.
As far as I could tell, this line of reasoning caused no distress among the scientists who found the hippo–whale link in their genetic data. Frankly, I'm not sure they knew what an astragalus was. How many computational biologists are trained in anatomy? The shape of the mesonychid astragulus was irrelevant to them, really. To them, DNA was an overwhelmingly superior source of information. It could be analyzed precisely. It was strings of code, rather than a maddening blur of phenotype.
I decided that this potential whale–hippo link was important enough to mention in my book, but did so only briefly because the results were so preliminary. Still, I left the matter open, and I'm glad I did. Subsequent studies on mammal DNA continued to support the whale–hippo link. And meanwhile paleontologists discovered more fossils of ancient walking whales. In 2001 Phil Gingerich and his colleagues from the University of Michigan and from Pakistan described a skeleton of the seal-like species Rodhocetus. Among its bones, they found an astragulus—the first whale astragulus ever discovered. And lo and behold, the astragalus had a double-pulley shape [3]. The DNA, it seems, had been right all along.
This experience made a strong impression on me. I was struck by the divide between these two kinds of biologists. Each group had a profound confidence in their own sources of information, and an abiding skepticism about the other's. As I learned more about the history of modern biology, I realized that this rift did not begin in the 1990s. It was already present in the 1950s, as molecular biologists began championing their new science over more traditional ways of understanding life.
Harvard University's biology department was a microcosm of this conflict. James Watson, fresh from discovering the structure of DNA, breezed into the department in 1956 with revolution on his mind. “It was time to sweep beyond mere description of animals and plants and move into a new biology based on chemistry and physics,” as Watson's biographer, Victor McElheny, writes [4].
Needless to say, the Harvard naturalists were not happy. Edward O. Wilson, entomologist, ecologist, and sociobiologist, pushed back hard. “Watson, having risen to historic fame at an early age, became the Caligula of biology,” he writes in his autobiography, Naturalist. “It was foolish, we argued, to ignore principles and methodologies distinctive to the organism, population, and ecosystem, while waiting for a still formless and unproved molecular future” [5]. The struggle only ended when Harvard's biologists agreed to split their department in two.
Over the past fifty years, these factions of biologists have had a complex relationship. Some scientists have continued to carry out relatively traditional natural history work, with little need to delve into molecular (or computational) biology. Others have given little attention to natural history, focusing their efforts instead on deciphering the complexities of a membrane channel, or building new algorithms for identifying open reading frames. In some cases, biologists have bridged this divide, and the result has been a fruitful collaboration. But in other cases—such as the DNA studies on whales and hippos—one group moves into the other's traditional territory, sparking new conflict.
“That can't be right” is a refrain I've often heard when computational biologists report a controversial result. Along with the whale–hippo link, I can also recall the snorts that came with the first studies on human DNA suggesting that we can trace our ancestry to Africa 150,000 years ago. The picture is certainly more complex than the first “out of Africa” scenarios suggested, but the core of that idea has withstood further scrutiny. These days I sense a growing confidence in computational biologists, perhaps brought on by successes such as these. Many are no longer content merely to present an analysis of DNA and leave it at that. They use their findings to make sweeping statements about ecology and evolution.
One example of this new ambition was a paper published earlier this year on the evolution of cats [6]. The scientists offered a sweeping scenario for cat evolution, complete with migrations of cats out of Asia into the New World and back, along with the emergence of the major groups of felids, ranging from ocelots to bobcats to lions. The scientists based their scenario entirely on an analysis of cat DNA. They did not consider a single fossil of a cat, nor did they have a paleontologist expert on cats as a coauthor. Cat fossil experts inform me that fossils of true cats as old as 17 million years have been discovered in North America. The geneticists put the arrival of cats in North America at only 8 million years ago. Whether or not the DNA results are correct, it is striking that the report does not even mention the existence of fossils that do not fit the pattern.
Genomes unquestionably contain a rich store of information, but sometimes that information can be deceptive. Recently, two paleontologists, Philip Donoghue of the University of Bristol and Mark Purnell of the University of Leicester, described how this deception can arise in the study of vertebrate evolution [7]. A number of studies have suggested that vertebrates underwent several rounds of genome duplication since they diverged from their closest invertebrate ancestors. One of these duplications may have coincided with the emergence of jawed vertebrates (gnathostomes). This link has prompted some scientists to propose that this duplication triggered a leap in complexity that set gnathostomes apart from their closest vertebrate relatives, the hagfish and lampreys. Gnathostomes not only have jaws, but also have a long list of other traits that set them apart from agnathans, including an advanced immune system, a mineralized skeleton, teeth, and fins. They are also far more diverse. Perhaps a genome duplication provided the raw material for an adaptive radiation.
As Donoghue and Purnell demonstrate, this idea only makes sense if you ignore the fossil record. Paleontologists have found a rich record of fossils documenting the gradual assembly of the gnathostome body plan. Conodonts, for example, possessed a mineralized skeleton, dentine, and enamel—but nothing else that sets gnathostomes apart from agnathans. Fins evolved later, and a mineralized braincase came later still. Nowhere along the phylogeny of vertebrates is there a sudden leap of complexity in the forerunners of true gnathostomes, nor is there an abrupt leap in levels of diversity. One is left to wonder just when the alleged effects of a genome duplication occurred.
Taking natural history seriously is good not only because it can steer a computational biologist away from these sorts of errors. It also opens up fascinating new questions to which computational biologists can profitably apply their skills. Consider the coelacanth, a rare, ugly fish found in the Indian Ocean. It belongs to the lobe fins, a group of fish from which we and all other land vertebrates evolved. Scientists knew about the fossils of coelacanths long before they saw one alive. The fossil record of their ancestors reaches back 400 million years, but it was not until 1938 that living coelacanths were discovered off the coast of South Africa.
I present the coelacanth in the spirit of the Buddha. Is it a fish, or a just bag of digital code?
Earlier this year, computational biologists published a remarkable paper about the coelacanth in Nature [8]. The paper had its origins in the startling discovery that certain regions of the human genome that do not code for proteins are highly conserved. Natural selection has kept these sequences much the same in humans and in other mammals. The scientists were puzzled as to how natural selection could preserve their homology, since these sequences do not encode any protein.
The scientists decided to see how widespread these regions are among animals. Searching gene databases, they discovered related genetic sequences in the coelacanth. Despite more than 400 million years separating our ancestors, the coelacanth and human regions were 80% identical. A close examination of the coelacanth segments revealed them to be transposons—virus-like stretches of DNA that could insert new copies of themselves into the genome.
The authors conclude that these ultraconserved regions of the human genome descend from a transposon in the ancestors of lobe fins. In the tetrapod lineages, the transposons lost their ability to spread through the genome. Some became exapted for regulatory functions, and were preserved by natural selection. The others were eroded by mutations. In the coelacanth lineage, on the other hand, the transposons remained active.
As fascinating as these results are, I was also struck by how they were framed. The authors begin their paper by referring to the coelacanth as a “living fossil.” They end their paper with the conclusion that by preserving this family of transposons, the fish “acts, in a sense, as a living molecular fossil. The remaining 99.9% of its genome, as yet unsequenced, may very well hold precious traces of additional events that helped shape our own evolution.”
The coelacanth is a living fossil only insofar as scientists once thought it was extinct. But that is not the dictionary definition of a living fossil: an organism “that has remained essentially unchanged from earlier geologic times” [9]. Paleontologists have been giving the coelacanth lineage a fresh look, and they've discovered a surprising amount of morphological change [10]. Some ancient relatives of today's coelacanths were slender, eel-like creatures. Others had round, sunfish-like bodies. Today's coelacanths are probably not living fossils in an ecological sense, either. Early fossils of coelacanths are often found in sedimentary rocks formed in coastal waters. Living coelacanths, by contrast, only live deep under water.
I would question whether the coelacanth is much of a molecular living fossil, either. The new research certainly shows that coelacanths preserve information in their genome that reveals some of the history of ultraconserved regions in our genome that would otherwise be lost. But that's because their transposons are still “alive,” while our transposons lost their ability to replicate a long time ago and have become frozen in place. In that sense, we are the living fossils.
If one thinks about the coelacanth's natural history as well as its genome, a new set of questions arises. Transposons are prone to mutations, which can take away their ability to insert new copies in a genome. These coelacanth transposons have been replicating for 400 million years, and now probably take up a significant fraction of the coelacanth genome. And yet they still have not diverged very much in all that time. Is natural selection conserving them? Is natural selection acting on the transposon or the coelacanth? How do these proliferating transposons affect the physiology of the coelacanth? Do they act like pathogens, or are they providing some benefit we don't yet understand? Does their expansion play any role in the ecology of living coelacanths?
None of these questions would even arise without the invaluable work of computational biologists. But if all the blind men gather together around the coelacanth, perhaps they can better understand just what this creature is.
The Buddha once told a story about a king who ordered a group of blind men to be presented with an elephant. Each man touched a different part of the animal. The king then asked them what an elephant is like.
The blind men who touched the elephant's head replied, “An elephant, your majesty, is just like a water jar.” The blind men who touched its ear said, “An elephant, your majesty, is just like a winnowing basket.” The blind men who touched its tusk declared, “An elephant, your majesty, is just like a plowshare.” The ones who touched the trunk replied, “An elephant, your majesty, is just like a plow pole.” The blind men who touched the body replied, “An elephant, your majesty, is just like a storeroom.” The blind men who touched the foot replied, “An elephant, your majesty, is just like a post.” The blind men who touched the hindquarters replied, “An elephant, your majesty, is just like a mortar.” The blind men who touched the tail replied, “An elephant, your majesty, is just like a pestle.” And the blind men who touched the tuft at the end of the tail replied, “An elephant, your majesty, is just like a broom.”
The blind men fell into a fistfight, shouting, “An elephant is like this, an elephant is not like that! An elephant is not like this, an elephant is like that!” [1]
I am a science writer, and my chief passion is biology. I spend time with biologists of all stripes—computational biologists, paleontologists, biochemists, ecologists, and all the rest. It is a marvelous privilege. But there are times, I must confess, when I feel like I am watching a blind fistfight.
One of the first bouts I witnessed took place in the late 1990s, when I was researching the origin of whales. Whales descend from terrestrial mammals, and made the transition to water between about 50 million and 40 million years ago. In the 1990s paleontologists began unearthing a series of spectacular fossils documenting that transition, including whales with full-blown legs. Functional morphologists joined forces to investigate the transition, studying swimming otters to understand how proto-whales might have moved through the water. I spent a lot of time with scientists such as these. They are naturalists. They have to know a lot of natural history. They have to memorize the details of many species, to understand how the physiology, behavior, morphology, and ecology of each animal add up into an integrated whole. Yet these naturalists also know that they only have a slippery grasp on all of that embodied complexity.
I put what I learned from those naturalists into my first book, At the Water's Edge [2]. As I was finishing up my manuscript, I began coming across papers in which scientists were taking a radically different approach to the question of whale origins: they were comparing the DNA of whales to that of other mammals.
At the time, molecular phylogenies were still a novelty. The computational methods for calculating them were relatively new, and scientists could only use them to compare a few gene sequences of whales and other mammals. But the conclusions from these few studies were the same: the closest living cousins of whales are hippos.
This may not seem like a big deal. It certainly didn't seem to bother the scientists who carried out the studies. They were just analyzing digital code, abstracted from the animals that carried it. The results spoke for themselves. Nevertheless, they gave paleontologists and mammalian systematists conniptions.
Here's why. Hippos are artiodactyls (also known as even-toed ungulates). Other artiodactyls include cows, camels, and pigs, and goats. Zoologists have long recognized a number of anatomical features that unite artiodactyls in their own group, distinct from other hoofed mammals such as horses and rhinos. One of the most obvious hallmarks was a bone in the ankle, the astragalus. The artiodactyl astragalus has a unique double-pulley shape that allows the artiodactyl hoof to swing back and forth in a distinctive way.
In the 1990s paleontologists found a number of spectacular early whale fossils, but they had yet to find bones from the whale ankle. The bones and teeth they did find suggested that the closest relatives of whales were an extinct group of mammals called mesonychids. Mesonychids were hoofed mammals, but they did not have a double-pulley astragalus. Therefore, paleontologists concluded, they were probably not artiodactyls. And if mesonychids were not artiodactyls, then whales could not be either. And that meant that the whale–hippo link had to be wrong.
As far as I could tell, this line of reasoning caused no distress among the scientists who found the hippo–whale link in their genetic data. Frankly, I'm not sure they knew what an astragalus was. How many computational biologists are trained in anatomy? The shape of the mesonychid astragulus was irrelevant to them, really. To them, DNA was an overwhelmingly superior source of information. It could be analyzed precisely. It was strings of code, rather than a maddening blur of phenotype.
I decided that this potential whale–hippo link was important enough to mention in my book, but did so only briefly because the results were so preliminary. Still, I left the matter open, and I'm glad I did. Subsequent studies on mammal DNA continued to support the whale–hippo link. And meanwhile paleontologists discovered more fossils of ancient walking whales. In 2001 Phil Gingerich and his colleagues from the University of Michigan and from Pakistan described a skeleton of the seal-like species Rodhocetus. Among its bones, they found an astragulus—the first whale astragulus ever discovered. And lo and behold, the astragalus had a double-pulley shape [3]. The DNA, it seems, had been right all along.
This experience made a strong impression on me. I was struck by the divide between these two kinds of biologists. Each group had a profound confidence in their own sources of information, and an abiding skepticism about the other's. As I learned more about the history of modern biology, I realized that this rift did not begin in the 1990s. It was already present in the 1950s, as molecular biologists began championing their new science over more traditional ways of understanding life.
Harvard University's biology department was a microcosm of this conflict. James Watson, fresh from discovering the structure of DNA, breezed into the department in 1956 with revolution on his mind. “It was time to sweep beyond mere description of animals and plants and move into a new biology based on chemistry and physics,” as Watson's biographer, Victor McElheny, writes [4].
Needless to say, the Harvard naturalists were not happy. Edward O. Wilson, entomologist, ecologist, and sociobiologist, pushed back hard. “Watson, having risen to historic fame at an early age, became the Caligula of biology,” he writes in his autobiography, Naturalist. “It was foolish, we argued, to ignore principles and methodologies distinctive to the organism, population, and ecosystem, while waiting for a still formless and unproved molecular future” [5]. The struggle only ended when Harvard's biologists agreed to split their department in two.
Over the past fifty years, these factions of biologists have had a complex relationship. Some scientists have continued to carry out relatively traditional natural history work, with little need to delve into molecular (or computational) biology. Others have given little attention to natural history, focusing their efforts instead on deciphering the complexities of a membrane channel, or building new algorithms for identifying open reading frames. In some cases, biologists have bridged this divide, and the result has been a fruitful collaboration. But in other cases—such as the DNA studies on whales and hippos—one group moves into the other's traditional territory, sparking new conflict.
“That can't be right” is a refrain I've often heard when computational biologists report a controversial result. Along with the whale–hippo link, I can also recall the snorts that came with the first studies on human DNA suggesting that we can trace our ancestry to Africa 150,000 years ago. The picture is certainly more complex than the first “out of Africa” scenarios suggested, but the core of that idea has withstood further scrutiny. These days I sense a growing confidence in computational biologists, perhaps brought on by successes such as these. Many are no longer content merely to present an analysis of DNA and leave it at that. They use their findings to make sweeping statements about ecology and evolution.
One example of this new ambition was a paper published earlier this year on the evolution of cats [6]. The scientists offered a sweeping scenario for cat evolution, complete with migrations of cats out of Asia into the New World and back, along with the emergence of the major groups of felids, ranging from ocelots to bobcats to lions. The scientists based their scenario entirely on an analysis of cat DNA. They did not consider a single fossil of a cat, nor did they have a paleontologist expert on cats as a coauthor. Cat fossil experts inform me that fossils of true cats as old as 17 million years have been discovered in North America. The geneticists put the arrival of cats in North America at only 8 million years ago. Whether or not the DNA results are correct, it is striking that the report does not even mention the existence of fossils that do not fit the pattern.
Genomes unquestionably contain a rich store of information, but sometimes that information can be deceptive. Recently, two paleontologists, Philip Donoghue of the University of Bristol and Mark Purnell of the University of Leicester, described how this deception can arise in the study of vertebrate evolution [7]. A number of studies have suggested that vertebrates underwent several rounds of genome duplication since they diverged from their closest invertebrate ancestors. One of these duplications may have coincided with the emergence of jawed vertebrates (gnathostomes). This link has prompted some scientists to propose that this duplication triggered a leap in complexity that set gnathostomes apart from their closest vertebrate relatives, the hagfish and lampreys. Gnathostomes not only have jaws, but also have a long list of other traits that set them apart from agnathans, including an advanced immune system, a mineralized skeleton, teeth, and fins. They are also far more diverse. Perhaps a genome duplication provided the raw material for an adaptive radiation.
As Donoghue and Purnell demonstrate, this idea only makes sense if you ignore the fossil record. Paleontologists have found a rich record of fossils documenting the gradual assembly of the gnathostome body plan. Conodonts, for example, possessed a mineralized skeleton, dentine, and enamel—but nothing else that sets gnathostomes apart from agnathans. Fins evolved later, and a mineralized braincase came later still. Nowhere along the phylogeny of vertebrates is there a sudden leap of complexity in the forerunners of true gnathostomes, nor is there an abrupt leap in levels of diversity. One is left to wonder just when the alleged effects of a genome duplication occurred.
Taking natural history seriously is good not only because it can steer a computational biologist away from these sorts of errors. It also opens up fascinating new questions to which computational biologists can profitably apply their skills. Consider the coelacanth, a rare, ugly fish found in the Indian Ocean. It belongs to the lobe fins, a group of fish from which we and all other land vertebrates evolved. Scientists knew about the fossils of coelacanths long before they saw one alive. The fossil record of their ancestors reaches back 400 million years, but it was not until 1938 that living coelacanths were discovered off the coast of South Africa.
I present the coelacanth in the spirit of the Buddha. Is it a fish, or a just bag of digital code?
Earlier this year, computational biologists published a remarkable paper about the coelacanth in Nature [8]. The paper had its origins in the startling discovery that certain regions of the human genome that do not code for proteins are highly conserved. Natural selection has kept these sequences much the same in humans and in other mammals. The scientists were puzzled as to how natural selection could preserve their homology, since these sequences do not encode any protein.
The scientists decided to see how widespread these regions are among animals. Searching gene databases, they discovered related genetic sequences in the coelacanth. Despite more than 400 million years separating our ancestors, the coelacanth and human regions were 80% identical. A close examination of the coelacanth segments revealed them to be transposons—virus-like stretches of DNA that could insert new copies of themselves into the genome.
The authors conclude that these ultraconserved regions of the human genome descend from a transposon in the ancestors of lobe fins. In the tetrapod lineages, the transposons lost their ability to spread through the genome. Some became exapted for regulatory functions, and were preserved by natural selection. The others were eroded by mutations. In the coelacanth lineage, on the other hand, the transposons remained active.
As fascinating as these results are, I was also struck by how they were framed. The authors begin their paper by referring to the coelacanth as a “living fossil.” They end their paper with the conclusion that by preserving this family of transposons, the fish “acts, in a sense, as a living molecular fossil. The remaining 99.9% of its genome, as yet unsequenced, may very well hold precious traces of additional events that helped shape our own evolution.”
The coelacanth is a living fossil only insofar as scientists once thought it was extinct. But that is not the dictionary definition of a living fossil: an organism “that has remained essentially unchanged from earlier geologic times” [9]. Paleontologists have been giving the coelacanth lineage a fresh look, and they've discovered a surprising amount of morphological change [10]. Some ancient relatives of today's coelacanths were slender, eel-like creatures. Others had round, sunfish-like bodies. Today's coelacanths are probably not living fossils in an ecological sense, either. Early fossils of coelacanths are often found in sedimentary rocks formed in coastal waters. Living coelacanths, by contrast, only live deep under water.
I would question whether the coelacanth is much of a molecular living fossil, either. The new research certainly shows that coelacanths preserve information in their genome that reveals some of the history of ultraconserved regions in our genome that would otherwise be lost. But that's because their transposons are still “alive,” while our transposons lost their ability to replicate a long time ago and have become frozen in place. In that sense, we are the living fossils.
If one thinks about the coelacanth's natural history as well as its genome, a new set of questions arises. Transposons are prone to mutations, which can take away their ability to insert new copies in a genome. These coelacanth transposons have been replicating for 400 million years, and now probably take up a significant fraction of the coelacanth genome. And yet they still have not diverged very much in all that time. Is natural selection conserving them? Is natural selection acting on the transposon or the coelacanth? How do these proliferating transposons affect the physiology of the coelacanth? Do they act like pathogens, or are they providing some benefit we don't yet understand? Does their expansion play any role in the ecology of living coelacanths?
None of these questions would even arise without the invaluable work of computational biologists. But if all the blind men gather together around the coelacanth, perhaps they can better understand just what this creature is.
Wednesday, March 5, 2008
An Exchange...
I recently had a discussion with a creationist at youtube. I think I've found the right method in dealing with them: Stay calm and rational, and after four or five responses, wish them the best, tell them you'll have to agree to disagree. Recommending a book never hurts either. I always try and recommend Ken Miller's "Finding Darwin's God"; or even better, a christian evolutionist website like Answers in Creation.
The Guy made several claims: The geologic column doesn't exist in its entirety anywhere in the world, bent strata indicate the rock was laid down catastrophically, and so on and so on. What bothered me is that he accused talk origins of making false claims, and insisted that there really was no complete geological column in one location. Luckily, talk origins always provides enough information for me to be able to check out their claims from other (and often better) sources. Sure enough, it does exist in one place. It disappoints me that creationists will swallow so much malarky from one or two creationist sources without bothering to do real research to get the facts.
Anyway, at one point we discussed human fossils. I asked why it was that humans are absent from all but the very top of the geologic column. He directed me here. There are several things about this article that are very hypocritical and downright stupid:
The first issue to consider is what we actually find in the fossil record.
~95% of all fossils are shallow marine organisms, such as corals and shellfish.
~95% of the remaining 5% are algae and plants.
~95% of the remaining 0.25% are invertebrates, including insects.
The remaining 0.0125% are vertebrates, mostly fish. (95% of land vertebrates consist of less than one bone, and 95% of mammal fossils are from the Ice Age after the Flood.)1
The number of dinosaur fossils is actually relatively small, compared to other types of creatures. Since the Flood was a marine catastrophe, we would expect marine fossils to be dominant in the fossil record. And that is the case.
Actually, sediments are usually deposited in water, so it isn't any surprise that most of the fossils we find are marine organisms. We certainly don't need Noah's flood to account for it! Another thing about this that is totally inconsistent is that this guy makes a huge deal out of only a few vertebrate fossils existing, yet creationists are constantly demanding finely graded chains fossils leading from past organisms to present (on occasion we actually find these, but they are rare and of course this only happens under the right conditions).
The article goes on to make speculations and invent excuses about why no human fossils exist in deeper strata. I stick to my original objection: It makes no sense for humans to have missed fossilization for the first 3 or 4 billion years of the fossil record, and then to have started fossilizing by the thousands around the Pliocene. That is, unless they didn't exist back then.
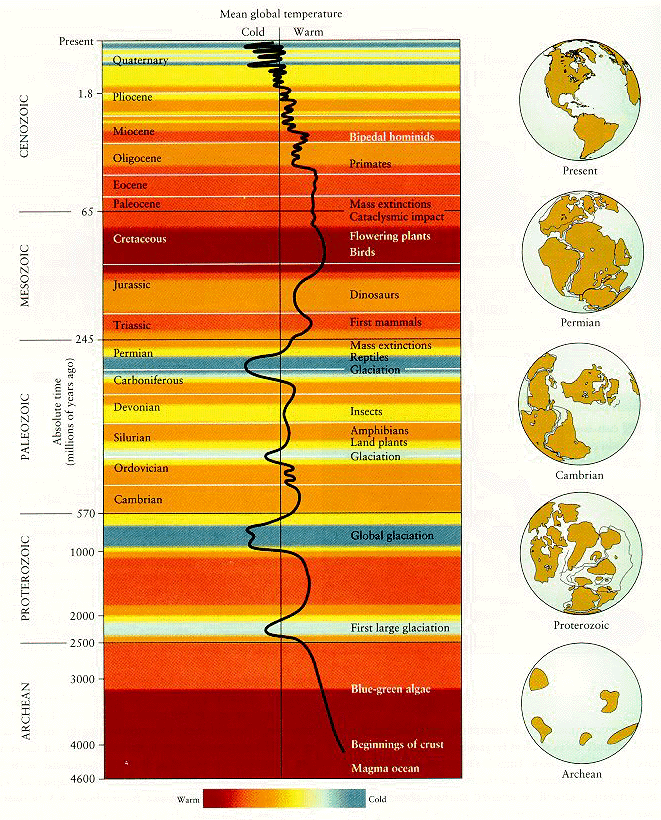
By the way, check out this link for a hilarious creationist take on the geologic timescale.
The Guy made several claims: The geologic column doesn't exist in its entirety anywhere in the world, bent strata indicate the rock was laid down catastrophically, and so on and so on. What bothered me is that he accused talk origins of making false claims, and insisted that there really was no complete geological column in one location. Luckily, talk origins always provides enough information for me to be able to check out their claims from other (and often better) sources. Sure enough, it does exist in one place. It disappoints me that creationists will swallow so much malarky from one or two creationist sources without bothering to do real research to get the facts.
Anyway, at one point we discussed human fossils. I asked why it was that humans are absent from all but the very top of the geologic column. He directed me here. There are several things about this article that are very hypocritical and downright stupid:
The first issue to consider is what we actually find in the fossil record.
~95% of all fossils are shallow marine organisms, such as corals and shellfish.
~95% of the remaining 5% are algae and plants.
~95% of the remaining 0.25% are invertebrates, including insects.
The remaining 0.0125% are vertebrates, mostly fish. (95% of land vertebrates consist of less than one bone, and 95% of mammal fossils are from the Ice Age after the Flood.)1
The number of dinosaur fossils is actually relatively small, compared to other types of creatures. Since the Flood was a marine catastrophe, we would expect marine fossils to be dominant in the fossil record. And that is the case.
Actually, sediments are usually deposited in water, so it isn't any surprise that most of the fossils we find are marine organisms. We certainly don't need Noah's flood to account for it! Another thing about this that is totally inconsistent is that this guy makes a huge deal out of only a few vertebrate fossils existing, yet creationists are constantly demanding finely graded chains fossils leading from past organisms to present (on occasion we actually find these, but they are rare and of course this only happens under the right conditions).
The article goes on to make speculations and invent excuses about why no human fossils exist in deeper strata. I stick to my original objection: It makes no sense for humans to have missed fossilization for the first 3 or 4 billion years of the fossil record, and then to have started fossilizing by the thousands around the Pliocene. That is, unless they didn't exist back then.
{kind=link}
By the way, check out this link for a hilarious creationist take on the geologic timescale.
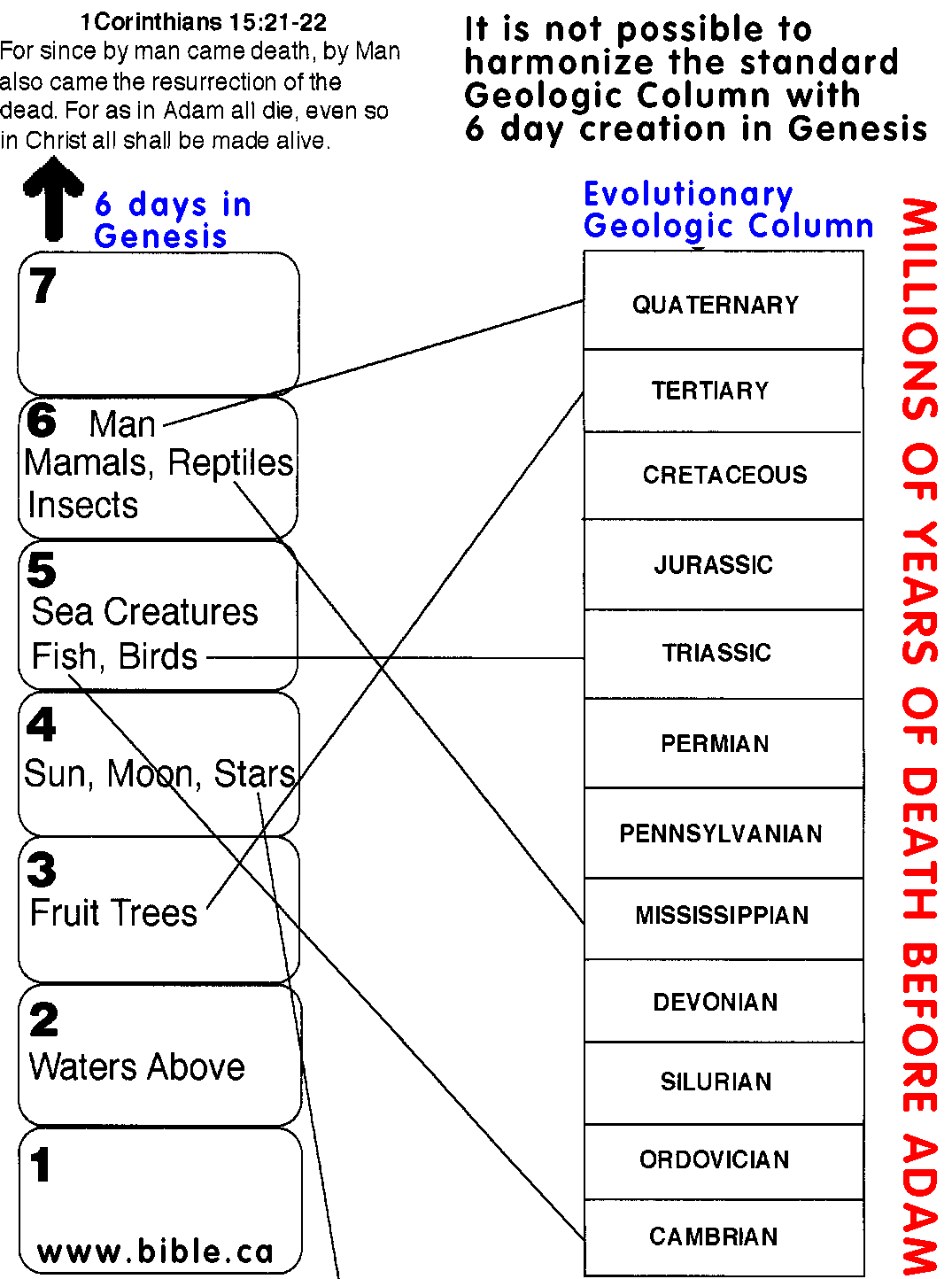{kind=link}
Sunday, March 2, 2008
Plausible Accounts of Abiogenesis
It is my intention in this post to illustrate the most feasible theories of how life began.
Scenario 1: The Clay Hypothesis
A.G. Cairns-Smith suggests that the first replicating beings were clay crystals. He argues that information is stored in the structure of the crystal, and passed on from generation to generation. Along the way, of course, the clay crystals who have structures that best allow them to reproduce eventually out compete the 'less fit' crystals. Cairns-Smith argues that the structure of the crystal could very well affect the chemistry of the environment it is in. He thinks that in the beginning some crystals were structured so that amino acids or sugars stuck to them. These crystals were benefited by their affinity for the molecules of life, and so the crystal population evolved this way (He speculates as to why and how sugars and amino acids would be useful to clay reproduction in his book, Seven Clues to the Origin of Life). Eventually, evolution worked its way up to clay crystals that produced nucleotides. These Nucleotides were later co-opted to serve as a new genetic system, and the primitive clay apparatus was dropped.
Pros and Cons:
This theory is very speculative and has little experimental evidence to support it. In fact, one recent study suggests that clay is not able to store genetic information very well. Now for the pros of this theory: if it is correct, the odds of life arising on earth would be 100%. Another pro is that it explains why we have not found any truly simple organism (Organic life outcompeted clay life long ago, and organic life by this time was relatively complex). To wrap up: I think a simple approach like this is needed for solving the mysteries of abiogenesis. Maybe Cairns-Smith's basic idea is right, except that clay wasn't the original replicator. Maybe some other self forming structure was.
For More, Visit this website:
http://originoflife.net/
Scenario 2: The Primordial Soup Theory
Scientists have been very successful in creating amino acids, nucleic acids, and even sugars under conditions that simulate the early earth (My Physical Science book cites an experiment by Carl Sagan that produced all of these). These experiments have been criticized for using atmospheres of methane and ammonia, which are currently not thought to be abundant in the atmosphere of the prebiotic earth. However, at least a few scientists are arguing that the atmosphere was mainly composed of methane, ammonia, and water vapor, just like the Miller Urey experiment used. Another point to be made is that many local areas, such as those with a lot of volcanic activity, would have had a reducing atmosphere. And finally, even if the prebiotic atmosphere was completely composed of carbon dioxide, hydrogen, and nitrogen, there are experiments that have used this atmosphere and produced amino acids.
Onto the theory: Scientists think that these organic molecules began bonding together to produce bigger and more complex molecules. Eventually, a molecule was formed that could replicate itself, and it evolved into life.
Several ideas exist about how these long molecules formed. Was it in hydrothermal vents? Did they form in ice? Or did it happen in between sheets of Mica? Several ideas exist about what the first replicator was, too. Was it a self replicating peptide, or an RNA molecule? And just how did this replicator evolve into life? This video has the most plausible illustration I have seen so far.
There are lots of less popular (and in my opinion, less plausible) theories on wikipedia. Discover Magazine has an article from a 1995 issue about "protocells" that David Deamer created. That about covers the origin of life theories. Yep.
Scenario 1: The Clay Hypothesis
A.G. Cairns-Smith suggests that the first replicating beings were clay crystals. He argues that information is stored in the structure of the crystal, and passed on from generation to generation. Along the way, of course, the clay crystals who have structures that best allow them to reproduce eventually out compete the 'less fit' crystals. Cairns-Smith argues that the structure of the crystal could very well affect the chemistry of the environment it is in. He thinks that in the beginning some crystals were structured so that amino acids or sugars stuck to them. These crystals were benefited by their affinity for the molecules of life, and so the crystal population evolved this way (He speculates as to why and how sugars and amino acids would be useful to clay reproduction in his book, Seven Clues to the Origin of Life). Eventually, evolution worked its way up to clay crystals that produced nucleotides. These Nucleotides were later co-opted to serve as a new genetic system, and the primitive clay apparatus was dropped.
Pros and Cons:
This theory is very speculative and has little experimental evidence to support it. In fact, one recent study suggests that clay is not able to store genetic information very well. Now for the pros of this theory: if it is correct, the odds of life arising on earth would be 100%. Another pro is that it explains why we have not found any truly simple organism (Organic life outcompeted clay life long ago, and organic life by this time was relatively complex). To wrap up: I think a simple approach like this is needed for solving the mysteries of abiogenesis. Maybe Cairns-Smith's basic idea is right, except that clay wasn't the original replicator. Maybe some other self forming structure was.
For More, Visit this website:
http://originoflife.net/
Scenario 2: The Primordial Soup Theory
Scientists have been very successful in creating amino acids, nucleic acids, and even sugars under conditions that simulate the early earth (My Physical Science book cites an experiment by Carl Sagan that produced all of these). These experiments have been criticized for using atmospheres of methane and ammonia, which are currently not thought to be abundant in the atmosphere of the prebiotic earth. However, at least a few scientists are arguing that the atmosphere was mainly composed of methane, ammonia, and water vapor, just like the Miller Urey experiment used. Another point to be made is that many local areas, such as those with a lot of volcanic activity, would have had a reducing atmosphere. And finally, even if the prebiotic atmosphere was completely composed of carbon dioxide, hydrogen, and nitrogen, there are experiments that have used this atmosphere and produced amino acids.
Onto the theory: Scientists think that these organic molecules began bonding together to produce bigger and more complex molecules. Eventually, a molecule was formed that could replicate itself, and it evolved into life.
Several ideas exist about how these long molecules formed. Was it in hydrothermal vents? Did they form in ice? Or did it happen in between sheets of Mica? Several ideas exist about what the first replicator was, too. Was it a self replicating peptide, or an RNA molecule? And just how did this replicator evolve into life? This video has the most plausible illustration I have seen so far.
There are lots of less popular (and in my opinion, less plausible) theories on wikipedia. Discover Magazine has an article from a 1995 issue about "protocells" that David Deamer created. That about covers the origin of life theories. Yep.
Subscribe to:
Posts (Atom)